Sự đề kháng Sulfamethoxazole của Streptococcus nhóm A phụ thuộc vào vật chủ
NTTU – Nhiều cơ chế đề kháng kháng sinh được các vi sinh vật sử dụng để đề kháng nhiều loại kháng sinh như bất hoạt kháng sinh bằng enzyme, giảm tích lũy kháng sinh, thay đổi đích tác động của kháng sinh hoặc bảo vệ đích tác động của kháng sinh trên vi khuẩn… Một cơ chế đề kháng kháng sinh mới đã được công bố trên tạp chí Nature Communications liên quan trực tiếp đến liên cầu khuẩn Streptococcus nhóm A có thể hấp thu folate dạng khử từ vật chủ để đề kháng kháng sinh Sulfamethoxazole.
Đề kháng kháng sinh (AMR – Antimicrobial resistance) là mối đe dọa rất lớn đối với sức khỏe con người trên toàn thế giới. Mỗi năm, chúng cướp đi sinh mạng của gần 1,27 triệu người trên toàn thế giới [1] và ước tính đến năm 2050 sẽ gây ra cái chết cho khoảng 10 triệu người mỗi năm. Đồng thời ước tính làm giảm tổng sản phẩm quốc nội (GDP) khoảng 2 – 3,5% và có thể tiêu tốn nguồn tài chính thế giới lên tới 100 nghìn tỷ USD [2].

Đề kháng kháng sinh của vi khuẩn
Hiện nay, sáu mầm bệnh hàng đầu được cho là có khả năng gây tử vong cao trên bệnh nhân có liên quan đến AMR là Escherichia coli, Staphylococcus aureus, Klebsiella pneumoniae, Streptococcus pneumoniae, Acinetobacter baumannii và Pseudomonas aeruginosa. Trong đó, S. aureus kháng meticillin gây tử vong hơn 100.000 ca, các dạng đề kháng khác như E. coli kháng cephalosporin thế hệ 3, A. baumannii kháng carbapenem, E. coli kháng fluoroquinolone, K. pneumoniae kháng carbapenem và cephalosporin thế hệ 3; mỗi loại gây tử vong khoảng 50.000 – 100.000 ca. Trong đó, các cơ chế đề kháng mà vi khuẩn sử dụng chủ yếu liên quan đến bất hoạt kháng sinh bằng enzyme, giảm tích lũy kháng sinh trong vi khuẩn, thay đổi biểu hiện của đích tác động hoặc bảo vệ đích tác động.
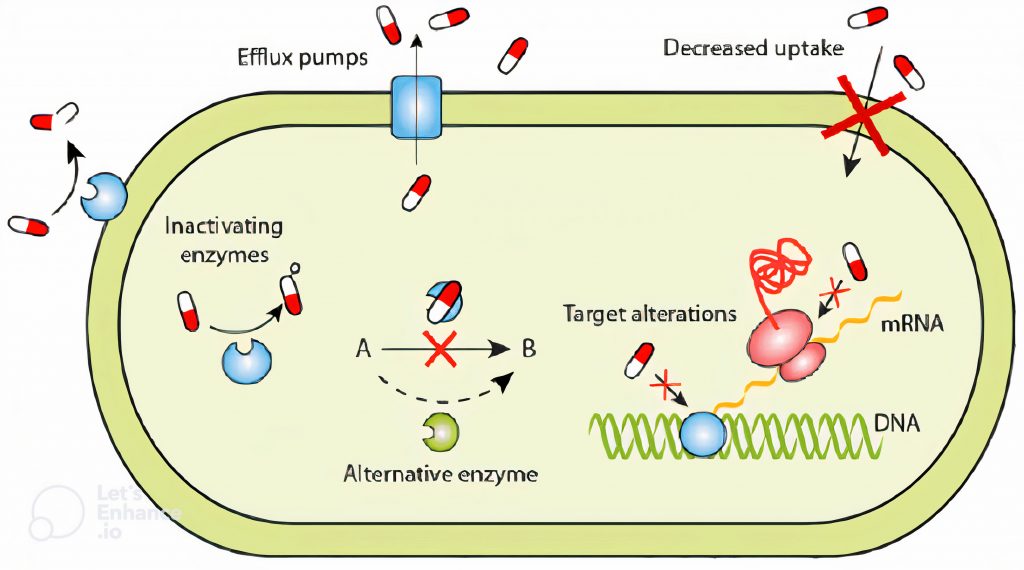
Các cơ chế đề kháng kháng sinh của vi khuẩn (theo E. Gullberg (2014))
Trong tự nhiên, tất cả các tế bào sống đều cần có folic acid (Vitamin B9) làm nguyên liệu cần thiết cho quá trình sinh tổng hợp deoxythymidine monophosphate (dTMP), một trong những loại nucleotide cần thiết cho tế bào để tổng hợp DNA khi nhân đôi tế bào. Vi khuẩn có khả năng tự tổng hợp folic acid để tồn tại tại và phát triển; trong khi, động vật có vú thiếu các enzym cần thiết cho quá trình sinh tổng hợp folic acid nên chúng phải thu nhận các folate dạng khử từ chu trình folate trong các nguồn thức ăn thu nhận. Các sản phẩm cuối cùng của chu trình folate là 7,8 – dihydrofolate – DHF và 5,6,7,8 – tetrahydrofolate – THF được sử dụng để tổng hợp deoxythymidine monophosphate (dTMP). Việc ức chế con đường tổng hợp folic acid có thể ức chế sự phát triển của vi khuẩn. Do đó, các liệu pháp kháng sự tổng hợp folic acid trong tế bào vi khuẩn đang được sử dụng phổ biến, trong đó có sự kết hợp hiệu lực cao hai loại kháng sinh đồng ức chế quá trình tổng hợp folic acid (Sulfamethoxazole – SMX) và chu trình folate (Trimethoprim – TMP). Sự kết hợp này (co – trimoxazole, SXT) thường được sử dụng để điều trị nhiều loại bệnh nhiễm trùng bao gồm cả S. aureus kháng methicillin liên quan đến cộng đồng, vi khuẩn Gram (-) đường tiết niệu và nhiễm trùng viêm phổi do Pneumocystis. SXT còn được khuyến nghị để điều trị nhiễm trùng da do Streptococcus pyogenes (Streptococcus nhóm A) ở các vùng lưu hành bệnh [3].
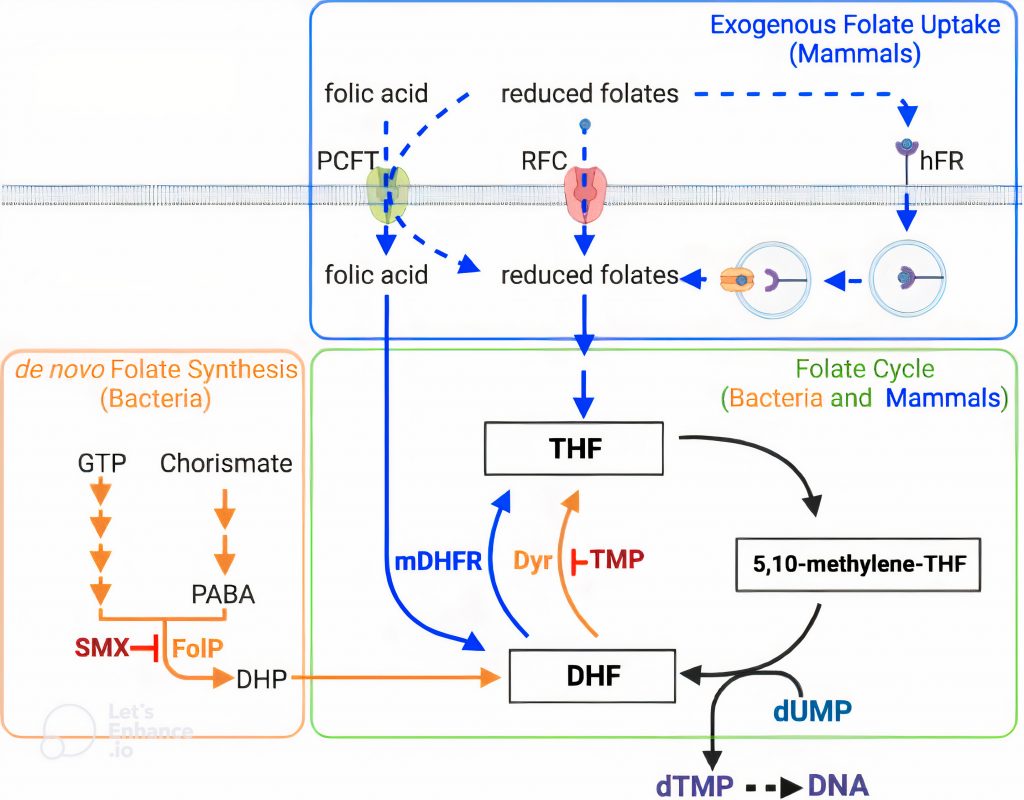
Tổng quan về chuyển hóa folate ở vi khuẩn và động vật có vú, và các đích tác động của SMX và TMP *
(* Dihydrofolate reductase của động vật (mDHFR) có thể khử folic acid thành DHF, các dihydrofolate reductase của vi khuẩn không có tác động. RFC và hFR là transporter cho folate dạng khử ngoại sinh, PCFT là transporter cho folic acid và folate dạng khử. RFC hoạt động chính trong các mô hệ thống, hFR và PCFT đặc hiệu cho mô.
RFC: reduced folate carrier, hFR: human folate receptor, PCFT: proton coupled folate transporter, GTP: guanosine triphosphate, DHP: dihydropteroate, DHF: dihydrofolate, THF: tetrahydrofolate, PABA: para-aminobenzoic acid, dUMP: deoxyuridine monophosphate, dTMP: deoxythymidine monophosphate, TMP: trimethoprim, SMX: sulfamethoxazole, DHFR: dihydrofolate reductase)
Mới đây, các nhà khoa học Australia đã xác định được một cơ chế đề kháng kháng sinh mới liên quan trực tiếp đến liên cầu khuẩn Streptococcus nhóm A – loại vi khuẩn gây nhiều bệnh nhiễm trùng từ viêm họng đến nhiễm trùng da. Theo kết quả nghiên cứu, khi Streptococcus nhóm A bị ức chế quá trình tổng hợp folic acid nội sinh bằng kháng sinh Sulfamethoxazole chúng vẫn có thể sử dụng folate dạng khử từ cơ thể vật chủ để phát triển, dẫn đến kháng sinh này không còn hiệu quả trên vi khuẩn [3]. Trong nghiên cứu kết hợp các thí nghiệm tiến hóa in vitro và giải cứu trao đổi chất, các nhà khoa học đã xác định được gen thfT cho phép Streptococcus nhóm A thu nhận các hợp chất folate dạng khử ngoại bào. ThfT có khả năng mở rộng tính đặc hiệu cơ chất của yếu tố vận chuyển ECF – transporter (Energy-coupling factor transporter) nội sinh để thu nhận các hợp chất folate dạng khử trực tiếp từ vật chủ, nhờ vậy mà vi khuẩn không bị ảnh hưởng bởi lượng folate nội sinh do chính vi khuẩn tạo ra. Khi không bổ sung folate dạng khử vào môi trường nuôi cấy mà ức chế vi khuẩn bằng kháng sinh thì tính đề kháng của Streptococcus nhóm A không còn, và tính đề kháng này chỉ xuất hiện khi vi khuẩn đang gây nhiễm trên vật chủ [3].
Kết quả thí nghiệm này cho thấy ThfT có chức năng tương đương với các con đường hấp thu folate dạng khử của sinh vật nhân thực, làm cho Streptococcus nhóm A có khả năng đề kháng kháng sinh Sulfamethoxazole rất cao. Điều đặc biệt là các phương pháp xét nghiệm hiện nay không thể phát hiện được tính đề kháng kháng sinh của nhóm vi khuẩn này trong in vitro, cơ chế đề kháng chỉ thấy được khi vi khuẩn gây nhiễm trên vật chủ. Trong bối cảnh AMR ngày càng gia tăng, điều quan trọng là phải có các công cụ chẩn đoán mới có thể nhanh chóng phát hiện tình trạng kháng thuốc kháng sinh, bao gồm cả tình trạng kháng phụ thuộc vào vật chủ thì việc điều trị mới đem lại hiệu quả [4].
ThS. Nguyễn Trung Hiếu – Khoa KTXNYH (tổng hợp)
Tài liệu tham khảo